|
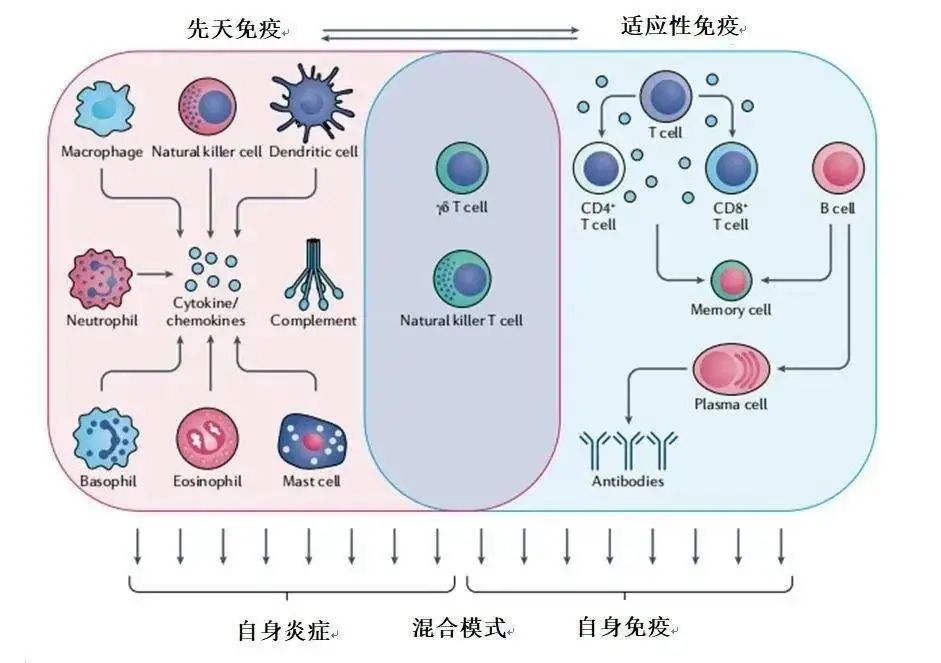
在许多人类自身免疫性疾病中,特别是在炎症的情况下,Treg细胞数量减少或功能受损。调节性T(Treg)细胞不能适当地控制免疫反应,必然导致自身免疫和器官损伤。白细胞介素2(IL-2)是驱动Treg细胞存活和功能的主要细胞因子,低剂量的IL-2恢复Treg细胞的适合度和/或扩大其数量,在早期临床试验中已显示出临床疗效。
IL-2的生物学:IL-2及其受体信号 IL-2是I型四α-螺旋束细胞因子,主要由传统T细胞在TCR和共刺激分子CD28参与后产生。激活CD4+T细胞和CD8+T细胞、NK细胞和NK T细胞、在某些情况下的树突状细胞和肥大细胞会产生IL-2,其中,CD4+T细胞所产生的水平最高。FOXP3会抑制IL-2在Treg细胞中的表达。Treg细胞并不能产生IL-2,而完全依赖IL-2的外源性供应来维持其生存和功能。
IL-2R由三个亚基组成:α链(IL-2Rα,CD25)、β链(IL-2Rβ,CD122)和γ链(IL-2Rγ,CD132)。IL-2R存在单体(IL-2Rα)、二聚体(IL-2Rβ - IL-2Rγ)或三聚体(IL-2-Rα-IL-2Rβ-IL-2Rγ)的三种变体,与IL-2的亲和力对应提高,Kd值分别为10−8M、10−9M、10−11M。
在IL-2结合中,下游信号通路主要有三个,分别是JAK-STAT(占总信号的90%)、PI3K-AKT-mTOR、和MAPK。IL-2结合导致IL-2Rβ和IL-2Rγ异二聚化,分别激活酪氨酸激酶JAK1和JAK3,使IL-2Rβ中的酪氨酸残基磷酸化,这促进了被JAKs磷酸化的信号分子如PI3K、STAT5或SHC1的募集,导致特异性途径激活、转录因子核易位,最终靶向转录调控,诱导细胞激活、分化和增殖。
IL-2的组成和信号通路
IL-2和Treg细胞、TH17细胞分化
IL-2是Treg细胞分化、免疫抑制功能、稳态和存活所必需的。Treg细胞控制着炎症反应,在炎症组织中变得效率较低,Treg细胞甚至可能因为失去FOXP3的表达而变得不稳定,并转变为一种更具有传统CD4+T细胞特征的表型,然后被称为“前T细胞”。同时,IL-2通过多种机制抑制TH17细胞分化。TH17细胞对于宿主抵御病原体至关重要,保护肠道免受炎症,但也参与了自身免疫性疾病的发病机制,如系统性红斑狼疮和牛皮癣。 因此,IL-2不仅对Treg细胞活性很重要,从而调节常规T细胞,也抑制TH17细胞的分化。
IL-2的临床应用
IL-2的临床应用于1984年首次被报道,以高剂量治疗刺激传统T细胞对肿瘤细胞的活性。FDA批准了高剂量IL-2用于治疗转移性肾细胞癌以及随后的转移性黑色素瘤。 在20世纪90年代早期,IL-2或IL-2R敲除小鼠表现出严重的自身免疫而不是预想的免疫缺陷。这种“IL-2悖论”可以用Treg细胞缺陷来解释。随后,IL-2在自身免疫中的作用的相关性变得清晰。2006年,研究人员推断Treg细胞组成表达高亲和力IL-2受体,在人类自身免疫性疾病中使用低剂量IL-2可能作为优先刺激Treg细胞活性而不激活效应T细胞的手段。事实证明确实如此,现在已经确定,在人体内每天300,000–3,000,000 IU的剂量下,IL-2主要刺激Treg细胞,并且耐受性良好。
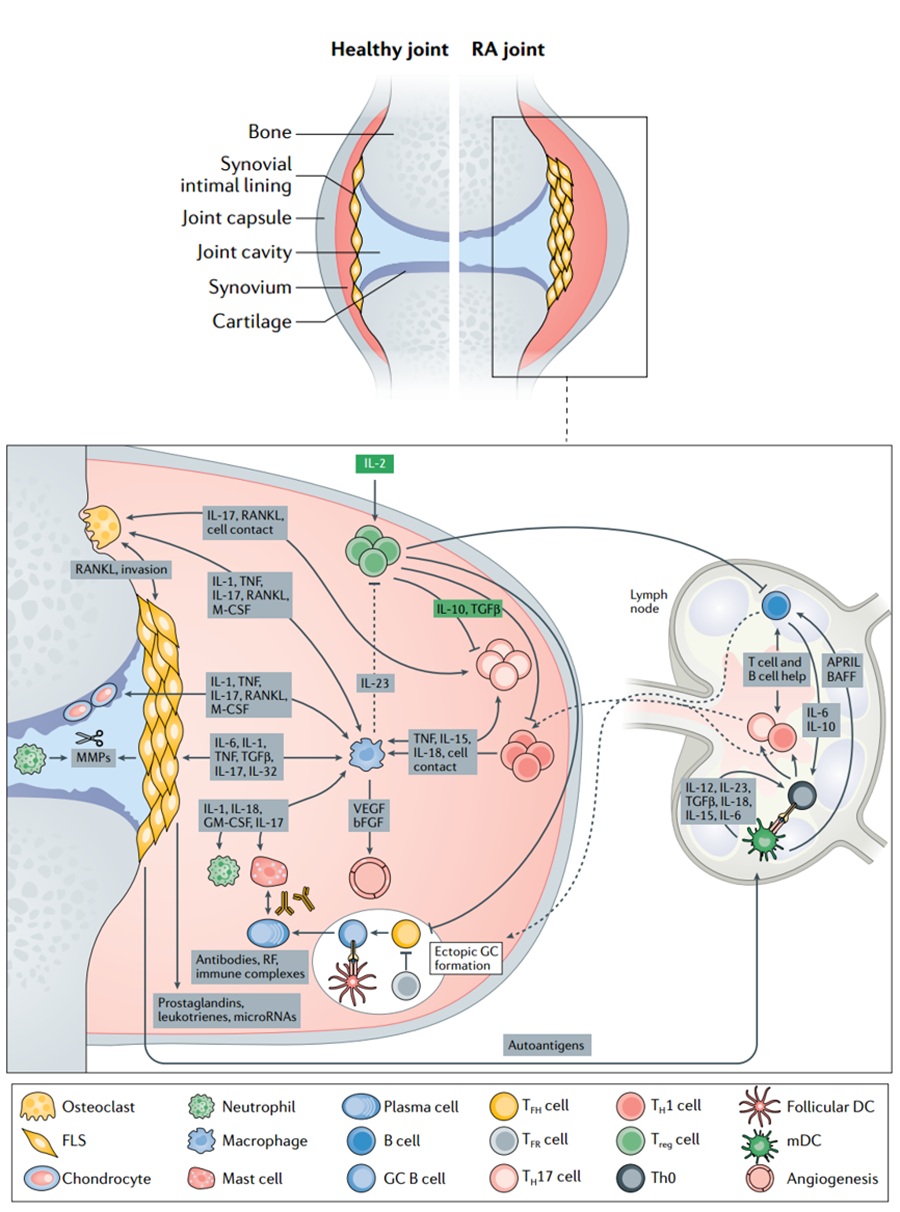
系统性红斑狼疮的发病机制 由于Treg细胞不足是大多数自身免疫性疾病发病机制的核心,对IL-2指标的检测和控制可能具有巨大的治疗潜力和广泛的临床适用性,如风湿性关节炎(三分之二的患者存在自身抗体,即血清阳性风湿性关节炎)、系统性红斑狼疮等。 |
Copyright © 2015-2023 杭州宇翼科技有限公司 丨 Discuz! X3.5 丨增值电信业务经营许可证:浙B2-20190572丨浙ICP备18026348号-1丨浙公网安备33010802009352号